|
|
|
|
|
|
Patrones de los peces de agua dulce en la vertiente
Caribe de Venezuela Consideraciones tomadas de: Rodríguez-Olarte, D., Taphorn, D. C., Lobon-Cervia,
J. 2009. Patterns of freshwater fishes of the Caribbean versant
of Venezuela. International Review of Hydrobiology, 91(1):67-90, Descargar.
Premio anual de investigación UCLA 2009. Suramérica contiene la mayor riqueza de
especies dulceacuícolas del planeta, presentando una mayor diversificación en
los grandes drenajes (Amazonas, Orinoco), pero también en ambos flancos de
los Andes y en las vertientes aisladas que drenan al mar. El estudio de la biogeografía de estas
faunas ha tenido un avance rápido y sus alcances actuales se expresan por la
revisión filogenética de grupos específicos, el estudio de la distribución
regional o continental; no obstante, algunas regiones o cuencas relativamente
pequeñas tienen historias geológicas que no necesariamente se asocian con los
patrones en la riqueza y distribución de especies reconocidos a mayor escala,
más aún, exponen procesos sólo a escala local. A parte de esto, la ictiofauna neotropical
aún es conocida parcialmente, principalmente por el variante conocimiento de
la sistemática para muchos taxa, insuficientes registros sobre su
distribución y dificultades en asociar las hipótesis filogenéticas con la
historia geológica, entre otras. Diferentes procesos geológicos han
propiciado la existencia de biotas particulares en las vertientes del Norte
de Suramérica, que a su vez han determinado regiones con faunas homogéneas (sensu lato provincias biogeográficas) poco
reconocidas, delimitadas o caracterizadas en su totalidad. Aparte de la
historia conocida para la región, se estima que varios procesos extinción y
especiación han ocurrido y permitido la actual riqueza de especies. En escalas geográficas amplias es previsible
discriminar dominios o provincias biogeográficas, tal y como es evidente en
la ictiofauna de la vertiente Caribe de Venezuela (VCV),
donde se reconocen inicialmente dos grandes biotas: Maracaibo y Caribe; sin embargo, este nivel de resolución impide
discriminar cuantitativamente la biota a menores escalas y limita el
reconocimiento de los procesos locales que inciden en el patrón biogeográfico
regional. Así, en escalas locales y regionales las biotas pueden tener una
heterogeneidad relevante, además de expresar gradientes particulares. Puesto que los eventos geológicos y
climáticos son conocidos en gran parte de la VCV es
posible relacionarlos con los datos sobre la riqueza y distribución de
especies para inferir sobre el actual patrón biogeográfico. En las cuencas de la VCV
no han sido reconocidos plenamente los patrones en la distribución de la
ictiofauna dulceacuícola que permitan identificar la variación faunística en
algunas regiones y menos aún como para identificar áreas de interés en la
conservación. En consecuencia, nuestro
objetivo fue dilucidar los patrones en la riqueza y distribución actual de
especies de peces dulceacuícolas en la VCV para
reconocer entidades biogeográficas y establecer su relación con procesos y
eventos geológicos y climáticos. Los métodos Atributos de las cuencas
hidrográficas.
Para efectos de este trabajo todos los drenajes al mar Caribe entre las
penínsulas de la Guajira y Paria componen la VCV. La agrupación y división de cuencas se
estableció con base en los mapas geológico y neotéctonico de Venezuela, así
como en los registros de relieve, superficie, altitud y divisorias de aguas.
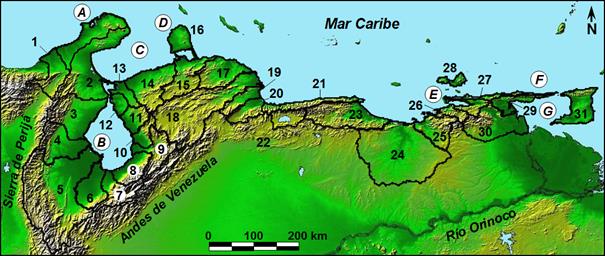
Las cuencas resultantes fueron (Figura 1): Limón, Palmar-Apón, Santa Ana,
Catatumbo, Escalante, Chama, Tucaní, Motatán, Misoa, Pueblo Viejo, Mene,
Cocuiza, Matícora, península de Paraguaná, Mitare, Hueque, Tocuyo, Aroa,
Yaracuy, Central, Valencia, Tuy, Unare, Neverí, Manzanares, Cariaco, isla de
Margarita, Paria, Ranchería (Guajira colombiana), San Juan y la isla de
Trinidad. Se consideraron drenajes
áridos y húmedos: la condición árida considera elevadas temperaturas y una
baja o muy baja pluviosidad. Las cuencas consideradas áridas fueron
Ranchería, Cocuiza, Matícora, Mitare, Paraguaná, Hueque, Ricoa, Tocuyo,
Unare, Neverí, Manzanares, Cariaco y Margarita, mientras que las restantes
fueron clasificadas como húmedas. Figura
1. Vertiente Caribe de
Venezuela y cuencas vecinas: (1) Ranchería, (2) Limón, (3) Palmar-Apón, (4) Santa
Ana, (5) Catatumbo, (6) Escalante, (7) Chama, (8) Tucaní, (9) Motatán, (10)
Misoa, (11) Pueblo Viejo, (12) Mene, (13) Cocuiza, (14) Matícora, (15)
Mitare, (16) península de Paraguaná, (17) Hueque y Ricoa, (18) Tocuyo, (19)
Aroa, (20) Yaracuy, (21) Costa central, (22) Lago de Valencia, (23) Tuy, (24)
Unare, (25) Neverí, (26) Manzanares, (27) Cariaco, (28) Margarita, (29)
Paria, (30) San Juan y (31) Trinidad Island. Hitos geográficos: Península de
la Guajira (A), Lago de Maracaibo (B), Golfo de Venezuela (C), Península de
Paraguaná (D), Península de Araya (E), Península de Paria (F) y Golfo de
Paria (G).
Bases de datos sobre la
ictiofauna. Se
seleccionaron 12155 registros de especies dulceacuícolas provenientes de las
colecciones de peces de CPUCLA, MCNG, MHNLS, EBRG y ICN-UNC y de las bases de datos de California Academy of Sciences y FishBase. Se dispuso de referencias generales para
actualizar los registros y se complementó la información con inventarios
locales y regionales. Se consideraron especies endémicas aquellas con
distribución restringida a una provincia o subprovincia dentro de la VCV. Las especies
con ocurrencia sólo en cuencas o subprovincias aledañas a la VCV (e.g. San Juan) se
consideraron restringidas, pues no se determinó su distribución general. Ya
que nuestro interés fue reconocer los patrones de distribución para la fauna
dulceacuícola estricta, no se consideraron aquellas especies o grupos
periféricos que ocurren en ambientes con influencia marina (desembocaduras,
estuarios) y aquellas con condición anfídroma, como las familias Gobiidae,
Ariidae y Gerreidae, entre muchas otras. Se elaboró una matriz conteniendo
registros binarios de presencia-ausencia para 34 familias y 270 especies
dulceacuícolas estrictas con ocurrencia en cuencas de la VCV,
Ranchería, Paria, San Juan y Trinidad.
Para reconocer los patrones de distribución según su tolerancia a la
salinidad se consideraron especies primarias y secundarias. La dispersión de
los peces primarios, fisiológicamente intolerantes a las condiciones salinas,
depende fundamentalmente por las conexiones entre drenajes, anastomosis,
capturas de cabeceras, eventos de precipitación o inundación extraordinarias,
mientras que los peces secundarios tienen tolerancia relativa al medio
salino, con lo cual podrían dispersarse a lo largo de los litorales marinos,
por lo que es considerado como un atributo que explique la presencia y
distribución de estas faunas a lo largo de la VCV. Clasificación y ordenación. Para clasificar y comparar las
relaciones de distribución para la ictiofauna se aplicaron análisis de
clasificación cluster mediante el algoritmo UPGMA empleando los coeficientes de similaridad de Jaccard y de disimilaridad
Euclidiana pues estos han sido efectivos para la resolución biogeográfica y
la comparación de los dendrogramas generados. Para probar agrupaciones
naturales en los datos se comparó cada matriz original con una matriz de
valor ultramétrico de los datos originales para obtener correlaciones
cofenéticas. Si dos matrices muestran
los mismos patrones de agrupación se produce una elevada correlación
cofenética, indicando una baja distorsión de los datos. Para contrastar los resultados de los
dendrogramas se desarrolló un análisis a escala multidimensional no métrica (NMS) basado en las mismas medidas de distancia. En el NMS la distorsión relativa de los datos originales fue
evaluada mediante los valores medios de stress; éstos decrecieron hasta
valores apropiados para representar la ordenación en el menor número de
dimensiones posibles. En los análisis multivariados no se incluyeron los
drenajes de la península de Paraguaná y la isla de Margarita pues no se
disponen de registros históricos adecuados y además contienen faunas
depauperadas. Con base en el análisis de riqueza y
distribución de especies y los análisis multivariados se detectaron las
entidades biogeográficas. En este trabajo se consideraron como dominios
biogeográficos a las regiones extensas, como los grandes drenajes regionales
o grupos de cuencas (e.g. lago de Maracaibo) con
muy pocas especies compartidas o muy baja similaridad, usualmente menor al
25%, respecto a otras cuencas o entidades. Las provincias representan
conjuntos de cuencas con biotas que tienen una similaridad de sus
faunas. Al nivel local, las
subprovincias se consideraron entidades menores, cuencas aisladas o pequeños
grupos de cuencas, con una elevada proporción de especies compartidas y
elevada similaridad. La designación de algunas entidades y sus límites
geográficos fue respaldada también por su ubicación espacial y su relación
con barreras geográficas evidentes.
Para inferir sobre los patrones detectados en la riqueza y
distribución de especies, las entidades biogeográficas fueron caracterizadas
en cuanto a su riqueza de especies, discriminando las de carácter endémico,
así como los eventos geológicos y climáticos reconocidos en la historia
pasada y reciente al norte de Suramérica. Resultados generales De las 34 familias evaluadas, los
Characidae (64 spp) y los Loricariidae (55 spp) abarcaron cerca del 45 % de
la riqueza general (ver Tabla 1: Lista
de especies de peces continentales en la vertiente Caribe de Venezuela, 2009).
Pocas familias sobrepasaron las diez especies, destacando los Cichlidae,
Rivulidae y Heptapteridae, entre otras; mientras que con cinco o menos
especies se reconocieron 25 familias. La mayor presencia de especies de peces
Characidae fue en las cuencas Tocuyo, Aroa y Tuy; mientras que los peces
Loricariidae fueron más especiados en las cuencas que drenan al lago de
Maracaibo. La riqueza de especies presentó un gradiente de cambio en sentido
de la longitud geográfica y con máximos valores en los drenajes al Lago de
Maracaibo y algunas cuencas particulares (e.g.
Aroa, San Juan). Las cuencas con mayor número de especies
(Catatumbo y San Juan) presentaron gradientes similares de especies que
comparten con otras cuencas de la VCV. La proporción de especies primarias (206
spp) fue generalmente elevada en cada cuenca (66%), mientras que la especies secundarias (64 spp) tuvieron promedios
menores, pero estos aumentaron hasta cerca del 50% en las cuencas de menor
tamaño y de condición árida, como en los drenajes al Norte del estado Falcón
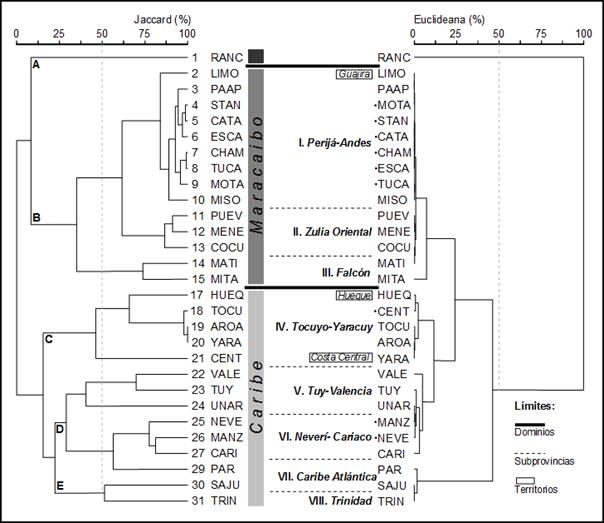
(e.g. Mitare). El dendrograma UPGMA
basado en la medida de distancia de Jaccard generó
un arreglo de las cuencas acorde con el geográfico espacial real (Figura 2). Los límites de VCV
correspondieron con los límites geográficos estimados inicialmente. Se reconocieron tres entidades con gran
diferencia entre sus biotas: Magdalena, Maracaibo y Caribe. Estas biotas tienen su división aproximada
entre los pares de cuencas de Ranchería-Limón, Mitare-Hueque y Cariaco-Paria.
La correlaciones cofenéticas para los dendrogramas basados en los coeficientes
de similaridad de Jaccard y de distancia Euclidiana
indicaron un elevado ajuste de los datos. En ambos dendrogramas se
identificaron grupos de cuencas con faunas homogéneas (Perijá-Andina, Zulia
Oriental, Mitare, Tocuyo-Yaracuy, Tuy-Valencia, Neverí-Cariaco, Caribe
Atlántica y Trinidad). De
igual manera, las ordenaciones NMS basadas en los
diferentes coeficientes discriminaron varias entidades para la ictiofauna. Figura
2. Relaciones generales entre las ictiofanas
en la VCV y cuencas vecinas. Los dendrogramas UPGMA son basados en los coeficientes de Jaccard y euclideo. Los límites
entre las entidades biogeográficas son representados por líneas punteadas.
Las barras verticales indican dominios. Las provincias son A: Magdalena, B:
Maracaibo, C: Caribe Occidental, D: Caribe Central y E: Caribe Oriental. Las
subprovincias son: Perijá-Andes (I), Zulia Oriental (II), Falcón (III),
Tocuyo-Yaracuy (IV), Tuy-Valencia (V), Neverí-Cariaco (VI), Caribe Atlántica
(VII) y Trinidad (VIII)].
Las diferencias a pequeña escala sugieren
que las biotas de peces en la VCV son heterogéneas
y pueden dividirse en entidades jerárquicas variadas. Algunos autores reconocen a mayores escalas
diferencias entre las biotas al Norte de Suramérica, separando las provincias
de Magdalena, Maracaibo, Costa de Venezuela y Trinidad y Tobago. En este estudio no se dispuso de
información que permitiese comprobar relaciones o límites biogeográficos a
escalas mayores (ejemplo: dominios); no obstante, las diferencias robustas
encontradas sugieren que a escalas regionales y locales los patrones
biogeográficos varían en gran medida y pueden ser decisivos en el
reconocimiento de entidades biogeográficas jerárquicas. Estos resultados han
permitido una regionalización en los patrones de distribución para la
ictiofauna a nivel local y regional en la vertiente Caribe de Venezuela. La comparación de las faunas entre varias
cuencas permitió reconocer los límites específicos y las relaciones con
drenajes al Caribe y Atlántico. Entidades
biogeográficas.
Dos dominios confluyen en la VCV: Maracaibo y
Caribe (Figura
3). El límite oriental del
dominio Magdalénico (cuenca Ranchería) y su ictiofauna tuvo muy baja relación
respecto a los otros dominios. La
diferencia en la contribución de especies compartidas entre las cuencas de
Limón (Maracaibo) y Ranchería, sugieren que esta última es parte de una
provincia dentro del dominio Magdalénico.
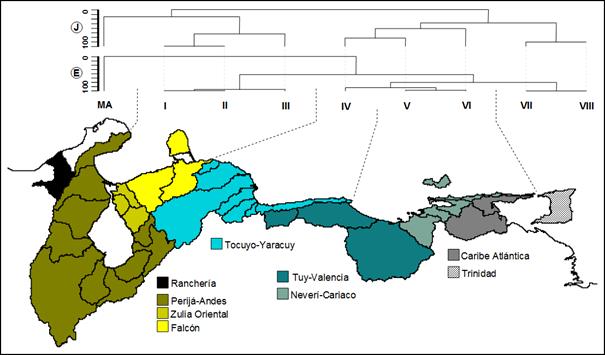
Cinco provincias biogeográficas son
reconocidas (Magdalénica, Maracaibo, Caribe occidental, Caribe central y
Caribe oriental) y presentan diferencias notables en el número de especies
endémicas (Figura
3). En las subprovincias detectadas (Ranchería, Perijá Andina,
Zulia Oriental, Mitare, Tocuyo Yaracuy, Tuy Valencia, Unare Cariaco, Caribe
Atlántica y Trinidad) se evidenció un gradiente de cambio de la riqueza de
especies, siendo la subprovincia Perijá Andina la más relevante en cuanto a
su endemismo y riqueza general de especies. La segunda subprovincia con mayor
riqueza y endemismo fue Tocuyo Yaracuy.
La subprovincia Caribe Atlántica, siendo
limítrofe con la VCV, presentó la mayor cantidad de
especies, pero muchas de éstas no necesariamente se restringen a las cuencas
de Paria y San Juan, siendo reportadas también en la cuenca del río
Orinoco. Algunas subprovincias
(ejemplo: Mitare) no presentaron ninguna especie endémica (sin embargo,
recientemente se describió un nuevo pez Loricariidae:
Ancistrus falconensis).
Por otro lado, la ocurrencia de taxa primarios y secundarios al nivel de
subprovincias mantuvo una proporción similar, registrándose mayor número de
especies secundarias en las subprovincias conformadas por cuencas áridas,
pero también de menor tamaño. En el texto original se describen de manera
extensa las subprovincias. Figura 3. Provincias y subprovincias detectadas
en la vertiente Caribe de Venezuela con base en los dendrogramas empleando
coeficientes de Jaccard (J) y euclídeo
(E). MA: Magdalena (dominio): cuenca de Ranchería.
Los refugios
hidrográficos. Se
postula que las variaciones climáticas durante los períodos de máximo e
intermedio glacial durante el Pleistoceno han incidido negativamente sobre
las biotas que se encuentran en los extremos de altitud, ya que estarían
expuestas a valores extremos de sequía y/o temperatura; caso contrario
ocurriría con aquellas poblaciones de peces asociadas a las vertientes
(piedemontes) donde las condiciones ambientales serían menos severas. Durante
el Pleistoceno los ambientes acuáticos podrían actuar como microclimas que
protegieron a la fauna de los extremos ambientales. Las cuencas de los ríos Aroa, Yaracuy y
Urama drenan un “abra geográfico” expuesto a los vientos alisios que aportan
humedad sostenida principalmente en las laderas y montañas, donde pueden
ocurrir precipitaciones orográficas. En el piedemonte de estas cuencas existe
la mayor riqueza de especies, incluyendo las endémicas, y por encima de los
800 msnm sólo dos especies son reconocidas.
Se estima que en momentos de máximo o
intermedio glacial las planicies aluviales y/o las máximas alturas locales en
la región (1500-2000 msnm) mantendrían condiciones extremas que limitarían la
permanencia de la ictiofauna dulceacuícola, pero las vertientes húmedas
servirían como refugios hidrográficos para el mantenimiento de la ictiofauna
local. Estos refugios hidrográficos se representarían como áreas aisladas en
el entorno regional menos húmedo. Un
refugio hidrográfico importante ocurriría en las planicies y vertientes al Sur
del lago de Maracaibo, donde ocurre la mayor riqueza de especies para esa
provincia. Esta región está sujeta a
una pluviosidad muy elevada ( Dada la historia geológica en la VCV, las áreas clave
detectadas también serían expresión de las perturbaciones de alcance regional
(e.g. trasgresiones, orogénesis) que permitieron la
alopatría, pero también de ambientes acuáticos heterogéneos y estables en las
vertientes de las montañas. La conformación geográfica de las áreas clave en
la VCV sugiere que los refugios hidrográficos no
estarían necesariamente reducidos al Pleistoceno sino que tendrían una
condición pretérita y una influencia constante hasta el día de hoy. Los refugios hidrográficos pueden ser
relacionados en parte con las hipótesis de Refugio y de Museo, las cuales
consideran que las poblaciones están sujetas a la especiación en ambientes
fragmentados o en sectores de alta montaña durante incursiones o ascensos del
nivel del mar en el Pleistoceno, para luego dispersarse a otros ambientes en
momentos climáticos menos restrictivos. Dispersión en la
vertiente Caribe de Venezuela.
Las especies primarias predominan en casi todas las cuencas estudiadas, pero
existen variaciones locales. La
ubicación de las cuencas con menor proporción de especies primarias puede
estar asociada a los ambientes con alguna influencia marina mientras las
condiciones regionales ancestrales afectaron negativamente el caudal de los
ríos en las cuencas altas y medias. En la VCV y sus
cuencas contiguas, los taxa con intolerancia a la salinidad se encuentran
principalmente en los Characiformes y Siluriformes (Tabla 1), con algunas
excepciones; mientras que los taxa secundarios son representados principalmente
por los Cichlidae, Poeciliidae, Rivulidae y Synbranchidae. La dispersión de los peces dulceacuícolas
en cuencas con planicies contiguas es reconocida, pero también a lo largo de
las costas marinas, ya sea por su condición de tolerancia a la salinidad o
por la reotaxia frente a las avenidas extraordinarias en los ríos, puesto que
las plumas generadas en la costa marina disminuyen la concentración de sales
en el agua, incluso a distancias considerables de las desembocaduras. Por otro lado, la dispersión también puede
estar asociada con la geomorfología terrestre y la plataforma submarina
contigua, propiciando una permeabilidad variable para la dispersión de
especies a través de las costas marinas.
Esto se evidencia en varios sectores donde las vertientes montañosas
se extienden hasta la costa y donde la batimetría litoral es somera (Figura 4).
En la VCV se distinguen tres sectores con estas
características: las estribaciones de la Sierra de San Luís en el estado
Falcón, las pequeñas cuencas de la cordillera de la Costa y las estribaciones
del macizo de Turimiquire y la península de Araya. En estos sectores el litoral es profundo,
al contrario de las playas someras que destacan para el resto de la VCV. Tales
características están asociadas con los cambios detectados en la riqueza y
composición de especies primarias y secundarias. Esto sugiere que la dispersión de muchos
taxa por vía marina puede estar limitada a determinadas secciones de la VCV, principalmente en aquellas cuencas contiguas (permeables)
y/o con fondos someros en sus desembocaduras al mar. Considerando que son
pocos los ríos con caudal importante que desembocan directamente al mar (ejempllo: Tocuyo, Tuy, Unare), la dispersión de la
ictiofauna por vía de las plumas fluviales (reotaxia) es más probable entre
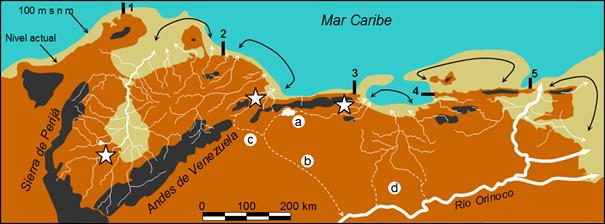
ríos con desembocaduras contiguas o cercanas y en costas de emersión someras. Figura
4. Procesos geofísicos que han estructurado la biogeografía en
la VCV. El nivel del mar, estimado en unos 120 por debajo
del actual hace unos 18000 años, permitiría conexiones entre los cursos bajos
de los ríos. Los registros sugieren posibles conexiones entre las cuencas de
Valencia y Tuy (a), y con la cuencas de los ríos Orinoco (b) y Yaracuy (c).
La dispersión (flechas negras) a lo largo de las costas pudo ser más intensa
en cuencas contiguas (Lago de Maracaibo) o entre desembocaduras de ríos y
deltas, pero limitada por la profundidad de la plataforma marina en algunos
sectores: Península de la Guajira (1), Norte costero de la subprovincia IV
(2), cuenca del territorio de la Costa Central (3) y los flancos de las
penínsulas de Araya y Paria (4 y 5). Las áreas claves se estiman como
refugios hidrográficos (estrellas) dentro de las subprovincias I, IV y V:
planicies al sur del Lago de Maracaibo, y cuencas de Aroa, Yaracuy y Tuy.
La ocurrencia de faunas de carácter
primario propias del Orinoco en drenajes al oriente de la VCV
puede tener relación con el enorme territorio ocupado por el delta de este
río, que colinda con la cuenca del río San Juan y tiene una influencia en el
golfo de Paria, pero igualmente puede estar asociada a la dispersión por
reotaxia en deriva gracias a la corriente ecuatorial del Norte y la baja
salinidad estacional promovida por el caudal del río Orinoco. El río Orinoco descarga al Atlántico
alrededor 31.000 m3/s y su pluma puede exceder los 160.000 km2;
superficie que cubre anualmente el Este y Sureste del mar Caribe. Al oriente
de la VCV y en los ríos contiguos, considerando la
dirección general de las corrientes marinas, los registros sugieren que sería
más frecuente la dispersión que el intercambio de especies, siendo un
fenómeno más intenso en las subprovincias Caribe Oriental y Trinidad. Sin embargo esta dispersión tiene poca
extensión en el resto de las cuencas de la VCV. En el pasado geológico y reciente el río
Orinoco funcionó como agente principal de dispersión en varios puntos de la VCV, pero los procesos regionales (ejemplo: aislamiento,
extinción, etc.) mermaron la permanencia de la biota orinoquense. Estas
diferencias en la distribución de la ictiofauna, reconocidas también entre
las entidades biogeográficas detectadas, serían expresión de las variaciones
en el nivel de mar en el pasado reciente.
En el Caribe, hace unos 18 Ka, el nivel del
mar alcanzaría alrededor de los -120 por debajo del actual (mbnm) y éste fue ascendiendo paulatinamente en el
Holoceno hasta el día de hoy. Las regresiones también fueron reconocidas en
el mar Caribe durante el Pleistoceno, donde alcanzarían, en un período
cercano al millón de años, alrededor de -45 mbnm. En las costas al norte de Suramérica
estas regresiones marinas permitirían la existencia de por lo menos un cauce
único final de los drenajes al lago de Maracaibo, lo que explica las especies
compartidas en las áreas bajas y medias de las cuencas. Igual ocurría con la
mayoría de las cuencas contiguas a lo largo de la VCV. Drenajes aislados, como la península de Paraguaná
y la isla de Margarita pudieron tener conexión con las cuencas
continentales. Para tales momentos las
cuencas que drenan al golfo de Paria, incluyendo las cuencas occidentales de
la isla de Trinidad, tendrían drenajes comunes. Así, en el pasado reciente, pudo existir un
intercambio importante de especies en varias cuencas de litorales de
Venezuela, siendo exitoso incluso para las especies primarias, pero el mismo
estaría supeditado posteriormente a la condición climática de las cuencas. |
|
|
|
©
Douglas Rodríguez Olarte. Colección Regional de Peces,
CPUCLA. Laboratorio de Ecología. Decanato
de Agronomía. Universidad Centroccidental Lisandro Alvarado. UCLA.
Barquisimeto, Lara, Venezuela. Primera
edición: Diciembre 2011. Última
actualización: Enero 2017. |
|