|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Biogeografía de los peces en el norte de Suramérica:
el legado del aislamiento, la extinción y la evolución en las cuencas de
Magdalena y Maracaibo Un extracto de: Rodríguez-Olarte,
D, Mojica, J. I., Taphorn D.C. 2011. Chapter Fifteen. Northern South America: Magdalena and Las cuencas de los ríos en el norte de
Suramérica (NSA) varían mucho en la composición
taxonómica de los peces continentales. Así, ríos con alta riqueza de especies y endemismo se
intercalan entre las regiones áridas con faunas empobrecidas (Dahl, 1971; Mago-Leccia, 1970). Estas
variaciones son producto de las actuales condiciones climáticas e
hidrológicas, pero también reflejan las transformaciones históricas sobre
estos drenajes. Acorde con la historia de NSA es conocido que el sistema proto-Orinoco-Amazonas
desembocaba en la cuenca del Caribe, un brazo occidental del Mar de Tetis. Con el
tiempo este sistema se fragmentó gracias a eventos tectónicos muy variados. En NSA la orogénesis de las diversas ramas de la cordillera
andina, que permitieron el nacimiento de las cordilleras Central y Oriental
de Colombia y los Andes venezolanos, y los movimientos de las placas
tectónicas asociadas, como las de América del Sur, el Caribe y la microplaca de Maracaibo, finalmente
dividieron las biotas acuáticas de manera permanente. Estas transformaciones
geológicas fueron enormes y acompañadas por importantes fluctuaciones en el
nivel del mar. Las incursiones marinas en el Plioceno temprano, cuando el
nivel del mar alcanzó a unos
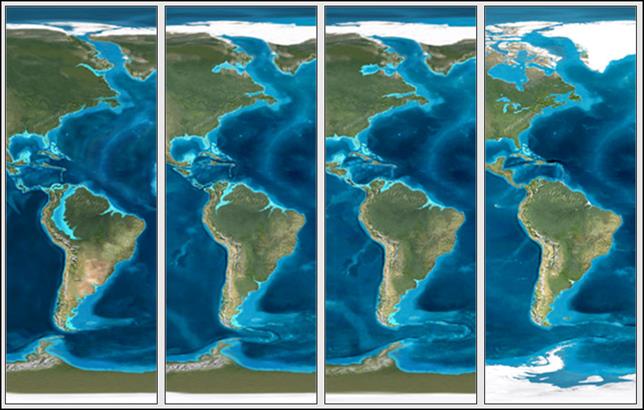
Se estima que los drenajes continentales al norte de
Suramérica han variado en gran medida a lo largo de las épocas geológicas, como
lo indica esta figura, que representa una secuencia desde el Eoceno hasta el
presente. Esto sugiere que los actuales patrones biogeográficos de las
ictiofaunas continentales podrían ser referencia a tales cambios. Esta figura
no está incluida en el artículo original (modificado de Global Paleogeography). Pero también hubo regresiones: por
ejemplo, se estima
que entre 20 y 18 mil años atrás (Ka), durante el
último máximo glacial, el nivel del mar descendió más de Análisis biogeográficos previos de los
peces continentales en NSA han mostrado escenarios
similares para el origen de las faunas de peces. Durante
la última mitad del siglo pasado, varios autores (ejemplo: Myers, 1966; Mago-Leccia, 1970; Taphorn y Lilyestrom, 1984; Galvis et al,
1997) reconocieron varias entidades zoogeográficas en NSA asociadas
con la riqueza de especies, los límites geográficos y la historia geológica. Los análisis posteriores se han concentrado en la
regionalización biogeográfica y el papel de la dispersión (Smith y Bermingham, 2005; Rodríguez-Olarte et al, 2009), el
análisis de parsimonia de endemismos (Hubert y Renno, 2006), el análisis filogenético utilizando el ADN
mitocondrial (Perdices et al., 2002) y la integración de los registros
biológicos con la historia geológica para explicar los acontecimientos
históricos (Lovejoy et al., 2006). En este trabajo
se presenta un examen de la riqueza de especies de peces continentales y sus
patrones de distribución aplicando técnicas de clasificación y ordenación
para delimitar unidades biogeográficas y, en consecuencia, relacionarlas con las
variables históricas y ecológicas. Los Métodos Selección
de cuencas y registros de las ictiofaunas: Como Norte de Suramérica (NSA)
se incluyen aquí los ríos costeros entre las cuenca
de Mira (Vertiente del Pacífico) y el Golfo de Paria (Vertientes del Caribe y
el Atlántico). Con
base en lo propuesto por Rodríguez-Olarte et al., (2009), en este trabajo
las cuencas cisandinas incluyen todas las cuencas al este de la península de
Paraguaná. Para
propósitos de comparación se incluyeron las vertientes Caribe y Pacífica de Baja
Mesoamérica (Smith y Birmingham, 2005), así como la cuenca del Orinoco como
una sola unidad biogeográfica. En la vertiente del Pacífico las cuencas escogidas fueron Mira, Patía, Dagua, San Juan (en adelante, San Juan Pacífico), Baudó y Tuira. En
la vertiente del Caribe las cuencas
fueron Chagres, Atrato, Sinú, Magdalena, Ranchería, Maracaibo, Cocuiza,
Matícora, Mitare, Hueque (incluyendo Ricoa), Tocuyo, Aroa, Yaracuy, Central
(que contiene varias cuencas muy pequeñas), Tuy, Valencia, Unare, Neverí,
Manzanares y Cariaco. En la
vertiente del Atlántico las cuencas escogidas fueron Paria (que agrupa pequeñas cuencas del Golfo de
Paria), San Juan (en adelante, San Juan Atlántico) y los ríos de la Isla de
Trinidad. Las
caracterizaciones, agrupaciones y
divisiones de las cuencas se establecieron a partir de bases de datos
variadas (CIET, 2005). Se utilizaron registros de
peces de agua dulce de las colecciones biológicas de CPUCLA, EBRG, MBUCV, MCNG y MHNLS. No se incluyeron
especies periféricas (por ejemplo, Gobiidae, Ariidae y Gerreidae). La
cobertura de las muestras de peces es amplia y suficiente para asumir que las
ausencias a nivel de cuencas son representativas. La riqueza de especies y su distribución: La riqueza y distribución de especies se analizó
considerando los niveles taxonómicos de familia, género y especie. Las especies se clasificaron en primarias y secundarias, en
función de su tolerancia a la salinidad (Stiassny y
Raminosoa, 1994). Las especies primarias (por ejemplo, Characiformes, Gymnotiformes),
no tienen o tienen muy baja tolerancia al agua salada, mientras que las
especies secundarias (por ejemplo, Cyprinodontiformes, Perciformes) son
tolerantes y tienen un mayor potencial para la dispersión a lo largo de
costas marinas. Para reconocer la variación en la riqueza de
especies respecto a diferentes modelos matemáticos, se desarrollaron modelos
de especies-área empleando funciones lineales y de potencia. Una curva de especies-área con alto valor residual
positivo sugiere que las cuencas tienen una riqueza superior a la media
predicha por el modelo (Fattorini, 2006). El mejor ajuste de los modelos se estimó
según el criterio corregido de
información de Akaike (AIC). Clasificación
y ordenación sobre unidades biogeográficas: Se elaboraron matrices de presencia y ausencia
para especies de peces estrictamente de aguas dulces o continentales (1391
especies). Para caracterizar y comparar las relaciones entre
las ictiofaunas se aplicaron análisis de conglomerados (clúster) usando el
algoritmo UPGMA y el coeficiente de similitud de Jaccard (Smith y Birmingham, 2005), empleando correlaciones cofenéticas como pruebas de bondad de ajuste (Rohlf y Fisher, 1968). Para
contrastar con la clasificación, se aplicó un análisis de escalamiento
multidimensional no métrico (NMS) con base en el
coeficiente de Jaccard. Las
coordenadas del NMS fueron generadas por un
análisis de correspondencia sin tendencia (DCA),
mientras que el coeficiente de determinación (r2) se aplicó como prueba de bondad de ajuste. Para
distinguir y caracterizar las entidades biogeográficas se analizaron los
valores de similitud obtenidos a partir de la clasificación multivariante y
los procedimientos de ordenación. Se
consideraron como especies
endémicas aquellas con distribución restringida a una sola provincia o
cuenca. En
este trabajo los
dominios fueron considerados como áreas muy extensas, tales como drenajes
regionales, grandes cuencas o grupos de cuencas (por ejemplo, la cuenca del
Orinoco) con faunas de peces homogénea que muestran similitud muy baja (por
lo general menos del 25%) respecto a las entidades vecinas. Las
provincias, en menor escala, representan grupos de cuenca con faunas que
tienen una similitud entre el 25-50%. Resultados generales La
riqueza y distribución de especies En NSA se
registran 511 especies primarias y secundarias, con 246 en las cuencas de
Magdalena y Maracaibo. En la vertiente Caribe las cuencas con mayor
riqueza de especies fueron Magdalena (159 spp., 66 endémicas), Atrato (120
spp., 19 endémicas) y Maracaibo (115 spp., 48 endémicas), todas ubicadas
en regiones húmedas. El resto de los ríos que desembocan en el Caribe o
el Atlántico mostraron una menor diversidad, con la excepción de unos pocos
originarios de la Cordillera de la Costa, como Aroa (59 spp.), Tuy (60 spp.)
y San Juan Atlántico (76 spp.). Las cuencas con mayor biodiversidad de peces
están separadas por pequeños ríos costeros con menor riqueza de especies, que
por lo general provienen de regiones áridas, como las costas del estado
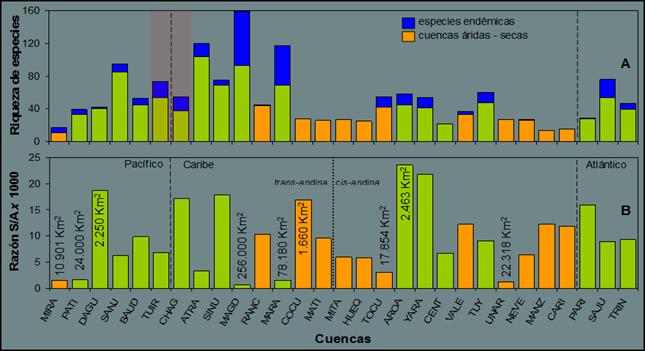
Falcón o la península de la Guajira, en Venezuela (Figura
1A). Figura 1. Variación de la riqueza especies de peces dulceacuícolas
primarias y secundarias en las cuencas de NSA,
excluyendo Orinoco. A) Variación en la riqueza según la secuencia geográfica
de los drenajes y B) la relación especie-área [(S / A) * 1000]. Las cuencas son: Mira (MIRA), Patía
(PATI), Dagua (DAGU), San Juan Pacífico (SANJ),
Baudó (BAUD), Tuira (TUIR), Chagres (CHAG), Atrato (ATRA), Sinú (SINU) , Magdalena (MAGD),
Ranchería (RANC), Maracaibo (MARA), Cocuiza (COCU), Maticora (MATI), Mitare (MITA), Hueque (HUEQ),
Tocuyo (TOCU), Aroa (AROA), Yaracuy (Yara) , Central (CENT), Tuy (TUY),
Valencia (VALE), Unare (UNAR), Neverí (NEVE), Manzanares (MANZ),
Cariaco (CARI), Paria (PARI), San Juan Atlántico (SAJU) y Trinidad ( TRIN). Las líneas discontinuas indican los
océanos Atlántico, Pacífico y el mar Caribe. Las barras grises indican las
cuencas secas y/o áridas. Nota: para 2010 fue descrita una especie endémica
para las cuencas de Hueque y Ricoa.
En las cuencas húmedas se registraron proporcionalmente más especies
primarias (~ 65%), que en las cuencas áridas, donde los peces secundarios
pueden alcanzar el 50% del la riqueza. Sin
embargo, existen excepciones como en los ríos Tocuyo y Unare, donde la
riqueza general es baja en relación con el tamaño de las cuencas
hidrográficas. La densidad de taxa por unidad de superficie mostró una
variación importante, pero la tendencia general es a disminuir en función del
aumento del área de drenaje: grandes cuencas tienen más especies y la
densidad por lo general es más baja respecto a las cuencas más
pequeñas. Las pequeñas cuencas de Aroa y Yaracuy tienen densidades muy
altas, con más de 20 especies por cada 1000 km2. Se determinó
que estas cuencas tuvieron la mayor riqueza de especies por unidad de
superficie. Así, estas cuencas abarcan unos 4.944 km2
(aproximadamente 0,9% del área total de estudio) y contienen alrededor de un
10,3% de todas las especies presentes en la NSA (Figura 1B).
El área de drenaje se correlaciona positivamente con el número de
especies presentes. El modelo de función lineal para la relación
especies-para las cuencas en NSA mostró un mejor
ajuste de la función de potencia (S
= 37,87 x 0,00083 (A); R² = 0,97);
donde S representa la riqueza de
especies, A representa el área de
la cuenca), mostrando un sólido diferencial de Akaike
(Δ AIC = 25,86;> 99,9 %). Cuando se
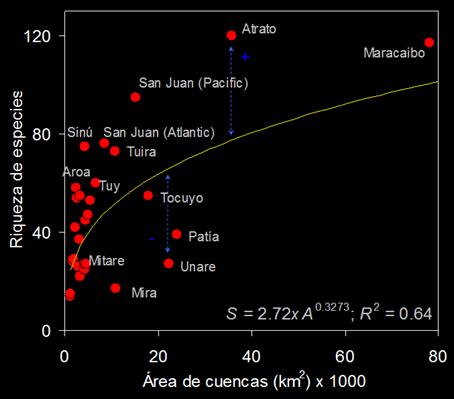
excluyó la cuenca del río Orinoco del modelo, éste produjo un mejor ajuste (S = 2,697 × (A) 0,3273; R² = 0,64; Δ AIC
= 8,32; 98,5%; Figura 2), aunque
su capacidad explicativa fue menor. El segundo modelo fue seleccionado
como la mejor representación de las relaciones especies-área en NSA. Las principales cuencas con mayor desviación de
los residuos positivos fueron Atrato, Sinú, San Juan (Atlántico y Pacífico) y
Aroa. Estas cuencas pueden ser consideradas con una elevada riqueza de
especies. La relaciones especie-área fueron sensibles a las cuencas con
riqueza de especies por debajo de la curva de ajuste. Estas cuencas se
caracterizan por tener climas secos o áridos. Figura 2. Relaciones especies-área registradas en una curva ajustada
mediante la función de potencia. La desviación de la media de este
modelo sugiere la existencia de drenajes con elevada o muy baja riqueza de
especies. La cuenca de Magdalena no se muestra por razones de
escala.
Las
familias y gradientes de diversidad
Las familias Characidae (157 spp.) y Loricariidae (102 spp.)
contribuyeron con casi la mitad (51%) de la riqueza total de especies en la NSA y fueron particularmente especiadas en las cuencas
más grandes, como Magdalena, Maracaibo y Atrato. Otras familias
importantes fueron Cichlidae y Trichomycteridae (30 spp;.
5,9%), Poeciliidae (26 spp), Heptapteridae (24 spp), Astroblepidae
(23 spp) y Rivulidae (23 spp; 4,5%). En conjunto, estas familias
constituyen cerca del 74% de todas las especies conocidas de la NSA. Para los Characidae el mayor número de especies
se registró en la cuenca de Magdalena (45 spp.), seguido por el Atrato (39
spp.), San Juan del Pacífico (24 spp.), Chagres (23 spp.), Sinú y del
Tuy, (22 spp. c/u). La familia Loricariidae alcanza su mayor número
de especies en la cuenca de Maracaibo (27 especies), la más alta de todas las
cuencas estudiadas de la NSA, tanto en cifras
absolutas y en porcentaje del total.
En NSA la riqueza de especies disminuye
hacia el este geográfico, pero aumenta de nuevo en los drenajes al oriente de
Venezuela, donde ocurren los géneros típicos del Orinoco (Crenicichla, Apistogramma y Astronotus). La
familia Trichomycteridae muestra su mayor diversidad en la cuenca de
Magdalena, con 11 especies en su mayoría endémicas. Los peces
eléctricos Gymnotiformes están expresados por las familias Apteronotidae (17
spp.) y Sternopygidae (12 spp.), mostrando una mayor diversidad en la cuenca
de Magdalena (9 spp.) y San Juan Pacífico (8 spp.). Clasificación
y ordenación
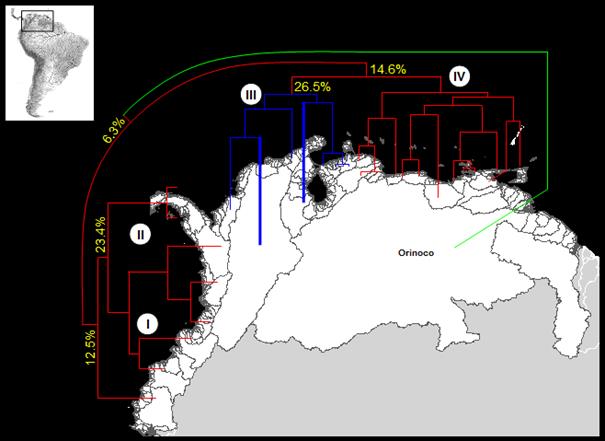
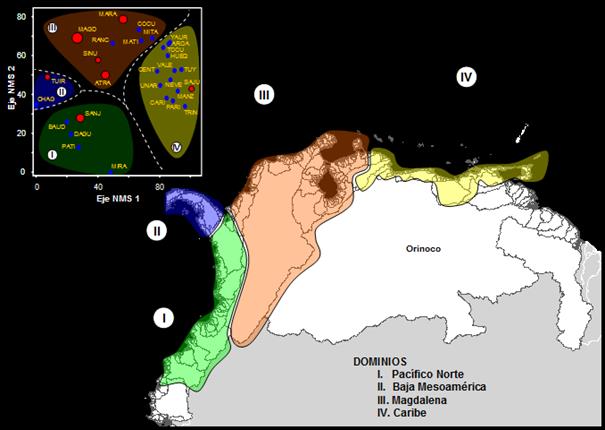
La disposición de las cuencas en NSA mostró
que el arreglo de los grupos generados fue similar al geográfico (Figura 3). Una alta correlación
cofenética (r = 0.91) indicó que los registros utilizados fueron adecuados.
Igualmente las ordenaciones NMS también se
mostraron adecuadas y sólidas (r2 = 0,58; ortogonalidad
= 98%; estrés = 20,1, Figura 4).
Las relaciones mostraron dos grandes grupos de faunas repartidas en NSA y Baja Mesoamérica. Estas subregiones tienen una
similitud muy baja en sus ictiofaunas. En el segundo nivel jerárquico y
con alta similitud se evidenciaron los dominios Pacífico Norte, Baja
Mesoamérica, Magdalena y Caribe.
Figura 3. Relaciones entre las especies primarias
y secundarias en la NSA y cuencas asociadas con
base en el análisis de clasificación según el algoritmo UPGMA
y el coeficiente de Jaccard (correlación cofenética
= 0.91). Las principales unidades biogeográficas
identificadas Pacífico Norte de Sur América (I), Baja Mesoamericano
(II), Magdalena (III) y el Caribe Norte de Sur América (IV).
Figura 4. Análisis multidimensional no
métrico (NMS) basado en el algoritmo UPGMA
y el coeficiente de Jaccard (r2 = 0,58; ortogonalidad = 98%; estrés = 20.1). La ordenación se ha rotado
por propósitos visuales. La disposición de todas las cuencas se relaciona con su
secuencia geográfica real. Los dominios biogeográficos se separaron: Pacífico (I),
Baja Mesoamérica (II), Magdalena (III) y el Caribe (IV). El tamaño de los símbolos
indica la magnitud de la riqueza de especies.
Los patrones biogeográficos emergentes de
los análisis demostraron la confluencia de cuatro grandes biotas en NSA asociadas con los océanos Atlántico y Pacífico, el
mar Caribe y Mesoamérica. En el ámbito del Caribe (103 especies, 46
endémicas), las tres provincias identificadas muestran diferencias
importantes: la Provincia Caribe Occidental tiene el mayor número de especies
endémicas (23,3%). La riqueza de especies para la provincia del Caribe
Central es la mayor y alcanza el 87% de todas las especies presentes en el
dominio, con el 16,1% de endemismo. Finalmente se identificaron y definieron
nueve provincias biogeográficas (Tabla
1): Patía (1), Atrato (2), Tuira (3 ), Chagres (4), Magdalena (5),
Maracaibo (6), Caribe Occidental (7), Caribe Central (8) y el Caribe Oriental
(9). Riqueza y distribución de especies y relaciones
especies-área La
distribución de los peces de agua dulce en NSA
muestra una correlación robusta con los principales aspectos geográficos y
climáticos de la región. La gran similitud de las faunas de peces las
cuencas de Magdalena y Maracaibo sugiere un origen común o por lo menos una
conexión ancestral entre ellos, que ha permitido la mezcla de sus peces, como
se ha señalado por diversos autores (Pérez y Taphorn, 1993; Albert
et al, 2006). El número de especies en común entre las cuencas de Magdalena,
Maracaibo y Orinoco difiere de la siguiente manera: (Magdalena-Maracaibo: 32
spp; Magdalena-Orinoco: 16 spp; Maracaibo-Orinoco: 21 spp), sugiriendo una
separación histórica gradual entre las mismas. Las cuencas de Magdalena y
Maracaibo comparten una mezcla de taxa que ocurren en diferentes altitudes;
pero muchas de estas especies conforman complejos taxonómicos con registros
en casi todos los drenajes costeros en NSA,
previéndose que futuras resoluciones taxonómicas disminuyan la similitud. Aún no
existe consenso sobre un modelo óptimo para las relaciones especies-área,
principalmente porque una única función no siempre refleja el mejor arreglo
biogeográfico o detecta patrones ecológicos (Scheiner,
2003), pero las aproximaciones son hoy una herramienta necesaria, como en
este caso. El aumento en la riqueza respecto al aumento de
la superficie de la cuenca es evidente como un parámetro general, pero hay
otros factores que afectan a esta relación. En NSA
el clima puede explicar muy bien por qué algunas cuencas de tamaño intermedio
tienen desviaciones importantes en sus relaciones especies-área. Esto se
podría asociar con cambios en el Pleistoceno como producto de desplazamiento
latitudinal de la Zona de Convergencia Intertropical y los cambios del nivel
del mar (González et al., 2008), que han provocado la desertización de
algunas cuencas y la extinción de sus ictiofaunas. Provincias biogeográficas En NSA la riqueza de especies y los patrones de distribución
se correlacionan con las modernas características orográficas, y en grado
variable, con la historia geológica y ecológica. En NSA las unidades biogeográficas designadas (Tabla 1) corresponden estrictamente a
las faunas de peces de agua dulce, extendiendo y discriminando unidades y
modelos previos que postularon que la ictiofauna en NSA
se distribuye en grandes provincias. Las unidades biogeográficas aquí
determinadas son: Provincias Patía y Atrato (Dominio Pacífico Norte
NSA): Se
identificaron dos provincias muy similares mediante análisis de similitud de
las faunas de peces en los ríos costeros del Pacífico colombiano (Figura 3). Una provincia está conformada por el río Atrato (vertiente Caribe) junto con los ríos Baudó y San Juan, mientras que la otra está formada por
los ríos Dagua y Patía. El
estado actual del conocimiento sobre la ictiofauna de esta región hace
difícil analizar el actual endemismo en estas cuencas. Provincias Chagres y Tuira (Dominio Baja
Mesoamérica): A pesar que las provincias
Chagres y Tuira tienen una similitud considerable respecto a la vertiente del
Pacífico en NSA, su ictiofauna pertenece a una
biota particular. Ambas provincias están estrechamente relacionadas con la
ictiofauna al norte de NSA. Un
consenso general sostiene que la NSA es la fuente
de los peces que colonizaron Mesoamérica gracias al levantamiento del istmo
de Panamá en el Plioceno y la creación de rutas de colonización (Myers,
1966). La ictiofauna de la provincia Tuira tiene una
menor similitud con la provincia de Atrato, lo que sugiere una especiación
rápida y/o extinción asociadas con la orogenia de los sistemas montañosos que
separan estas cuencas. Provincias
de Magdalena y Maracaibo (Dominio Magdalena): Las fuertes afinidades
ictiológicas entre las cuencas del Magdalena, Maracaibo y Orinoco han sido reconocidas
por varios autores (Pérez y Taphorn, 1993). Los fósiles de peces registrados en el valle del
Alto Magdalena (Arapaima, Colossoma, Lepidosiren, Phractocephalus) ratifican la
existencia de una biota ancestral que ocupaba un paleodrenaje
que hoy se ha dividido en las cuencas del Amazonas y el Orinoco (Lundberg et al., 1998). Este
tipo de patrón de distribución es también conocido por otros géneros y
familias de peces. Tabla 1. Unidades biogeográficas reconocidas en
la NSA y cuencas vecinas según los análisis de
clasificación y ordenación. Por cuencas: número total de familias (F),
géneros (G) y especies (S). A: la cuenca Mira está en la frontera del área de
estudio. B: En la actualidad drena en el mar Caribe. C: agrupa al a comarca Kuna Yala en Panamá. D: el dominio Orinoco es sugerido. Una nota para la página electrónica: Para
muchas cuencas, principalmente las aquí consideradas húmedas, el número de
especies ha aumentado ligeramente pero de manera constante debido a la
descripción de nuevas especies y la revisión de grupos.
Alrededor de un 21% de los peces de agua dulce en NSA
se registran en la provincia de Maracaibo. Algunas de sus especies
primarias tienen una distribución disyunta respecto a otras provincias, lo
que podría ser explicado por la historia geológica regional. Muchas
especies en el drenaje del río Limón (en el límite noroeste de la cuenca de
Maracaibo) tienen carácter endémico. El intercambio de las ictiofaunas
es evidente entre los ríos compartidos con planicies aluviales y esto ha
permitido la dispersión y la colonización entre los ríos de la cuenca de
Maracaibo. La mayoría de las especies registradas en las llanuras del
sur del lago (por ejemplo, Pterygoplichthys, Perrunichthys,
Platysilurus) están asociadas con
hábitats complejos de las planicies aluviales.
La proporción de especies endémicas característica de estas planicies
de inundación en las tierras bajas alcanza el 70% y sólo unas pocas especies
endémicas están restringidas a grandes alturas (por ejemplo, Astroblepus). La provincia
Maracaibo contiene una alta proporción de especies primarias (75%). Aquí
la riqueza disminuye hacia el noreste y en la costa del estado Falcón,
cambiando la proporción de especies con el predominio de las
secundarias. Los drenajes costeros de Falcón tienen faunas empobrecidas
con una mezcla de especies de los dominios Magdalena y Caribe. La
distribución de la ictiofauna y las condiciones climáticas actuales y pasadas
indican que las cuencas áridas de la provincia de Maracaibo han sido
colonizadas por especies de los ríos que drenan directamente en el lago. Provincias
Caribe Occidental, Central y Oriental (Dominio Caribe NSA):
El límite
oriental del dominio Magdalena es evidente (c. 5% de similitud) en los
drenajes áridos del estado Falcón (Figura 3), donde existe una sustitución
radical de las especies que componen
la ictiofauna. Dos áreas de endemismo son notorias: Aroa-Yaracuy y Tuy,
que en conjunto aportan el mayor porcentaje de la riqueza de especies y
endemismo de la provincia. En este dominio Rodríguez-Olarte et al. (2009)
reconocieron tres provincias (Caribe Occidental, Central y Oriental). La
provincia Caribe Occidental tiene la mayor riqueza de especies (72 spp.), así
como el mayor endemismo (23 spp; 32%).
La cuenca del río Tocuyo, con su origen en los flancos andinos, carece
de ciertos grupos, como los bagres Astroblepidae
familia común en las vertientes llanera y al lago de Maracaibo de los Andes
venezolanos (Maldonado-Campo et al., 2005). Este hiato es aparentemente
relacionado con las barreras geográficas, las condiciones climáticas y la
extinción. Así, en las cuencas de Aroa y Yaracuy se encuentran especies
de otras provincias e incluso especies de la cuenca del Orinoco, pero además
en el área tan pequeña de esas cuencas existe una elevada riqueza de especies
y endemismo. Esto se puede explicar por el aislamiento y la existencia de
refugios hidrográficos en las estribaciones de las montañas. Este
aislamiento ha fomentado un rápido proceso de especiación vicariante,
expresada en varios linajes (Characidae, Loricariidae, etc.).
En la provincia Caribe Central, la separación de baja altitud de
algunas cuencas (ej. Unare), así como la deposición de sedimentos
provenientes del río Orinoco sugiere una conexión de estos drenajes en el
pasado. El Lago de Valencia tiene especies compartidas con cuencas vecinas. La
separación de la cuenca del Lago de Valencia de la del Tuy se estima en el
Pleistoceno. Algunas especies son reconocidas como endémicas de la
cuenca del Valencia (por ejemplo, Atherinella venezuelae, Lithogenes valencia,
Pimelodella tapatapae),
pero otras, como Moenkhausia pittieri, también se han encontrado en la cuenca del
Tuy, lo que indica una conexión de estas cuencas en el pasado
reciente. La baja diversidad de especies en esta cuenca endorreica puede
explicarse por la inestabilidad en un clima con predominio histórico de la
aridez.
En la provincia Caribe Oriental los ríos Neverí, Manzanares y Cariaco
tienen la menor riqueza de especies. A pesar que Serrasalmus neveriensis se reporta como
endémica del río Neverí, otros registros sugieren que su mayor presencia y
distribución ocurre en el río Tuy, indicando que la especie ha colonizado
diferentes cuencas contiguas. La mayor parte de estas ictiofaunas están
asociadas con las del Orinoco; además, la cuenca de San Juan Atlántico
muestra biotas asociadas a los drenajes del Atlántico. La fauna de peces
de la Isla de Trinidad, por ejemplo, comparte el 60% de sus especies con las
de ríos que drenan al Golfo de Paria, lo que también sugiere conexiones
ancestrales, principalmente por las regresiones marinas y la confluencia de
ríos locales.
La composición de especies en esta provincia biogeográfica sugiere una
dispersión y colonización reciente por parte de la ictiofauna entre cuencas
costeras de diferentes provincias. A pesar que el Delta del Orinoco ha sido y
sigue siendo un núcleo permanente de la dispersión de los peces a lo largo de
la costas orientales de la NSA, la intensidad de su
efecto es variable debido a la dilución de las aguas dulces acorde con
cambios en la profundidad de la plataforma continental, lo que ha limitado la
dispersión de los peces de agua dulce en esta región. |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
©
Douglas Rodríguez Olarte. Colección Regional de Peces,
CPUCLA. Laboratorio de Ecología. Decanato
de Agronomía. Universidad Centroccidental Lisandro Alvarado. UCLA.
Barquisimeto, Lara, Venezuela. Primera
edición: Diciembre 2011. Última
actualización: Enero 2017. |
|